Here is the story of one unsung scientist in our field — who was likely the first person to identify ribosome hibernation factors, in their series of works between 1978 and 1982 (long before their rediscovery by Akira Wada and colleagues in 1990), but whose work never accrued sufficient visibility to reach a larger audience (or, in fact, any audience). Their name was García-Patrone, an Argentinian scientist from Buenos Aires.
For historical context, by the late 1970s, researchers studying ribosomes were puzzled by a long-standing mystery. Shortly after the discovery of ribosomes by Palade in 1953, many research labs found that ribosomes isolated from stationary phase cells have a low capacity for protein synthesis in a test tube, in vitro. For instance, Li & Umbreit (1966) showed that ribosomal fractions from E. coli cells in early log phase were far more active in synthesising polyphenylalanine than fractions from cells harvested only a few hours later. Later, González, Goldemberg & Algranati (1968) — scientists from the Fundación Campomar institute in Buenos Aires (the home institute for García-Patrone) — showed the same pattern in Bacillus stearothermophilus. For this organism, amino acid incorporation by ribosomes from late stationary phase cells was ten times lower than that of growing cells, suggesting that the low activity of stationary phase ribosomes was a broadly conserved phenomenon across bacteria.
Importantly, a few years later, Scheps, Wax & Revel (1971) found that this inactivity was not an intrinsic defect of the ribosomal particles themselves, because inactive ribosomes from stationary phase E. coli could be reactivated for translation in vitro by high-salt washing or puromycin treatment. This finding strongly suggested that something was bound to the ribosomes and suppressing their activity.
Parallel evidence came from a completely different biological system obtained by the Laboratory of Molecular Embryology at Rome. Metafora, Felicetti & Gambino (1971) showed that ribosomes isolated from unfertilised sea urchin eggs — which are translationally silent until fertilisation — were less active in polyphenylalanine synthesis than ribosomes from fertilised eggs, and that a high-salt wash of these ribosomes yielded a fraction that potently inhibited translation when added back to an active system. The inhibitory activity was destroyed by trypsin but not by RNases, suggesting the factor was proteinaceous in nature. Taken together, these studies painted an emerging picture that ribosomes in metabolically dormant cells are held silent by associated factors, likely proteins, that can be released by salt — though the identity of these factors remained unknown.
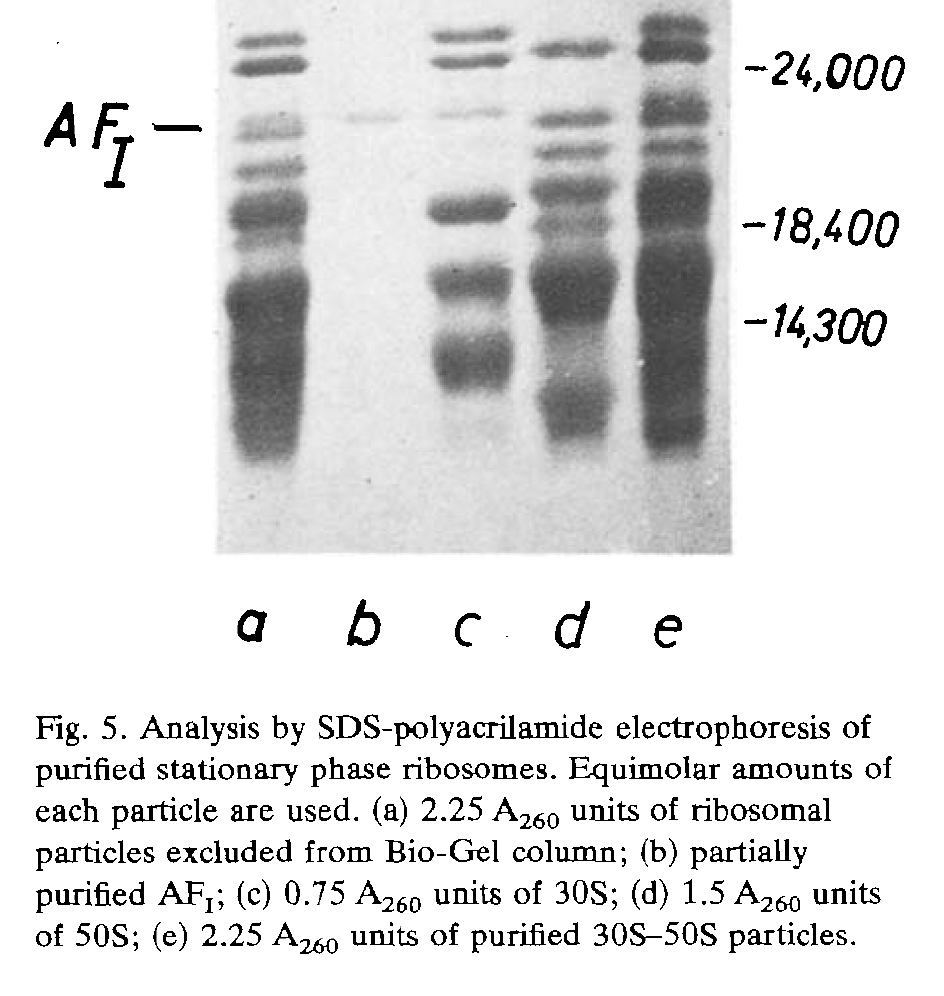
It was García-Patrone who first set out to identify one. Working with Bacillus stearothermophilus — the same thermophilic bacterium studied by their colleagues at the Fundación Campomar institute — García-Patrone showed that the drop in translational activity that accompanies entry into stationary phase could be attributed to a specific protein that accumulates on ribosomes as cells stop growing. This protein, named Association Factor I (AFI), was found on 30S subunits and on intact 70S ribosomes, but absent from 50S subunits alone — pointing to the small subunit as its binding site. Crucially, AFI was absent from mid-log phase ribosomes, appeared in late-log phase particles at low levels, and was present in abundance on stationary phase ribosomes — a pattern that tracked precisely with the decline in translational activity across the growth cycle.
Furthermore, when García-Patrone washed stationary phase ribosomes with high salt (which dissociates ribosomes from most ribosome-binding proteins and nucleic acids), AFI was quantitatively released and their activity was fully restored to levels comparable to those of actively growing cells, while the overall protein content of the ribosomes decreased by only 13%, confirming that AFI was not a core ribosomal protein but a loosely associated factor. Mechanistically, García-Patrone showed that AFI acts at the initiation step of translation: it could not inhibit polyphenylalanine synthesis once elongation had already started, it failed to bind ribosomes engaged in active protein synthesis or assembled into polysomes, however it blocked the binding of fMet-tRNA and mRNA to the ribosome. With a molecular weight of approximately 20 kDa, and given its association with the 30S subunit, its accumulation specifically during stationary phase, and its ability to silence ribosomes by blocking translation initiation, AFI almost certainly represents what we now know as HPF — the hibernation promoting factor — a protein that has since been identified across a wide range of bacteria and shown to play a central role in ribosome hibernation during dormancy.
Here is the kicker: despite this foundational discovery, most of which was described in the 1980 paper, thsi study was cited only once(!), by their own follow-up work from 1982(!). And it took the community another decade until Akira Wada and colleagues identified the ribosome hibernation factor RMF in E. coli, after which the study of ribosome hibernation factors slowly gathered momentum.
I personally deeply admire these early studies by a scientist almost no one knows about, and send my retrospective gratitude to this person — a pioneer who ventured into the unknown of one of the cell’s most remarkable superpowers: the ability to survive extended periods of starvation and return to life to keep the fire going.
References
- Li, L., and Umbreit, W.W. (1966). Relationship between ribosomal activity and age of culture in Escherichia coli B. Biochim. Biophys. Acta 119, 392–399.
- González, N.S., Goldemberg, S.H., and Algranati, I.D. (1968). Protein synthesis and ribosomal distribution at different growth stages in Bacillus stearothermophilus. Biochim. Biophys. Acta 166, 760–762.
- Metafora, S., Felicetti, L., and Gambino, R. (1971). The mechanism of protein synthesis activation after fertilization of sea urchin eggs. Proc. Natl. Acad. Sci. USA 68, 600–604.
- Scheps, R., Wax, R., and Revel, M. (1971). Reactivation in vitro of inactive ribosomes from stationary phase Escherichia coli. Biochim. Biophys. Acta 232, 140–150.
- García-Patrone, M. (1978). FEBS Lett. 92, 263–267.
- García-Patrone, M. (1980). Mechanism of protein synthesis inhibition during stationary phase in Bacillus stearothermophilus. Mol. Cell. Biochem. 31, 97–103.
- García-Patrone, M. (1982). The reactivation of stationary phase ribosomes from Bacillus stearothermophilus. Biochem. Biophys. Res. Commun. 104, 1413–1419.
- Wada, A., Yamazaki, Y., Fujita, N., and Ishihama, A. (1990). Structure and probable genetic location of a “ribosome modulation factor” associated with 100S ribosomes in stationary-phase Escherichia coli cells. Proc. Natl. Acad. Sci. USA 87, 2657–2661.